Generation Time
by Joachim
Gruber
The generation time of a single
cell is the duration TG of a cell cycle, measured as the
time between two consecutive mitoses.
In a bacteria population the single-cell
generation time varies during the development of the population. Each phase
of this evolution has its own single-cell generation time TG
or its specific generation time distribution TG, min ... TG,
max. When giving the single-cell generation time one has to specify
the phase of the development in which one has measured it.
One distinguishes the following phases
of the development (Stephen
T. Abedon, Important
words and concepts (3/10/01) from Chapter 6, Black, J. G. (1999). Microbiology.
Principles and Explorations. 4th ed., Prentice Hall, Ohio
State University, Mansfield, OH 44903, USA):
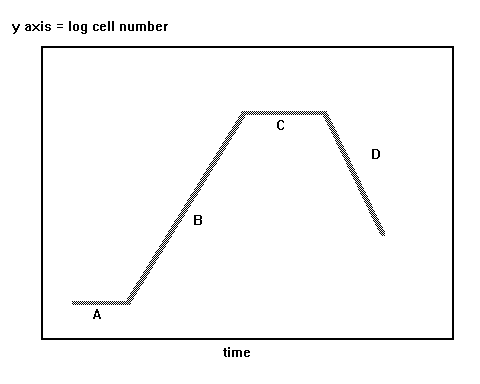
-
lag phase A, no cell diviaion. This
lag in division is associated with a physiological adaptation to the new
environment, by the cells, prior to their resumption of division. That
is, cells may increase in size during this time, but simply do not undergo
binary fission.
-
log phase B during which binary fission
occurs. This phase of growth is called logarithmic or exponential because
the rate of increase in cell number is a multiplicative function of cell
number. The generation time of the population (sensu stricto) is
the time it takes for the population in phase B to double in number of
bacteria.
-
Continuous culture phase C.
-
Decline (die-off, kill) phase.
Typically this die-off occurs exponentially in time. This death occurs
because vegetative cells can survive exposure to harsh conditions (few
nutrients, too many toxins, antibiotics) for only so long.
Microbiological literature reports the
kill kinetics of Borrelia burgdorferi. The borrelia population decreases
in these experiments exponentially with a half life
TS = 10 ... 12 hours dependent amog others from the antibiotic,
its time of action, Bb species and Bb genotype. (Abb. 1, Heinemann M et
al.).
-
Agger WA, Callister SM, Jobe DA,
In
vitro susceptibilities of Borrelia burgdorferi to five oral cephalosporins
and ceftriaxone, Antimicrob Agents Chemother 1992;Aug;36(8):1788-90.
-
Heinemann M, Trautmann M, Sepsis
und Antibiotika-induzierte Freisetzung von Endotoxinen: Konsequenzen für
die Therapie? Chemotherapie-Journal, 8. Jahrgang, Heft 5, 1999.
-
Preac-Mursic V, Marget W, Busch U,
Pleterski D, Rigler S, Hagl S , Kill
Kinetics of Borrelia burgdorferi, Infection, 24: 9 - 16, 1996.
As is explained in parentheses that
follow
-
the generation time TG (in
absence of the cell wall antibiotic) and
-
the kill kinetics half life TS
(in the presence of the cell wall antibiotic)
are of similar size.
(Here is an explanation for it:
In principle, the population can
be killed with the same rate as it is cell dividing (1.),
with a smaller rate (2.) or a larger rate
(3.):
-
If the cell wall antibiotic
TS = TG.
-
If the cell wall antibiotic does not
kill each cell that divides, some cells survive and thus the killing
(in the presence of the antibiotc) is slower than the cell division in
the absence of the antibiotic. In other words: TS is the upper
limit of the Bb generation time TG (i.e. TS > TG).
The Bb generation time is then smaller than the above mentioned 10
... 12 hours. This is consistent with the culture experiments by e.g.
Pollack et al.
Pollack RJ, Telford SR
3rd und Spielman A, Standardization
of medium for culturing Lyme disease spirochetes, J Clin Microbiol
1993,May;31(5):1251-5.
-
If the cell wall antibiotic kills the
cell that is dividing plus other cells, killing (in the presence of the
antibiotic) is faster than cell division (in the absence of the antibiotic):
TS < TG.
Interestingly, it has been
observed that in the presence of the antibiotic up to twice as many cells
die per unit time as would divide in the absence of the antibiotic. In
other words: 1/2 TG < TS < TG.
Koop AH, Neef C, van Gils SA.
A
mathematical model for the efficacy and toxicity of aminoglycoside (April
2003). Workshop "Predictive Value of PK/PD models of antimicrobial
drugs". Leiden University Medical Center, Leiden, The Netherlands, 4 -
5. September 2003.
Mouton JW, Vinks AA, Punt NC.
Pharmacokinetic-Pharmacodynamic
Modeling of activity of ceftazidime during continuous and intermittent
infusion. Antimicrobial Agents and Chemotherapy 1997;1(4):733-738.
Vinks AA, Punt NC and Mouton
JW. Pharmacokinetic-Pharmacodynamic
Modeling of Bacterial Growth and Killing using the Modified Zhi Emax model.
Workshop "Predictive Value of PK/PD models of antimicrobial drugs". Leiden
University Medical Center, Leiden, The Netherlands, 4 - 5. September 2003.
On the whole, TS and
TG have roughly the same size.)
Some
related basics in pharmakodynamics.
version: March 1, 2004
URL
of this page
Home
send comments to Joachim
Gruber